
by Ian St George
Sentences like “In vesicular-orbuscular mycorrhizas, the mycobiont undergoes pronounced alterations in morphogenesis involving appressoriuin formation, (and) arbuscule development…” leave the amateur gasping for breath. This is a complex subject with its own expert language: fortunately writers like Warcup and Perkins from Australia demonstrate the ability to convey complex information simply.
Fungi
What is a fungus? We think of toadstools, bracket fungi, ringworm, athlete’s foot – they are all fungi. The fungi are called Mycota, about 50,000 species described, including mushrooms, yeasts, rusts, smuts, mildews and molds. The mushroom we eat is the fruit (also known as the “perfect state”) of a larger organism, which has hollow branched filaments called hyphae that form networks in the soil; the bracket fungus we see is the fruit of a larger organism whose hyphae thread the dead bark and wood of the tree. Ringworm is the infection of the human skin by similar organisms; if you scrape the skin cells and look at them under the microscope you can see fungal hyphae.
The study of fungi is mycology, and many words referring to fungi have “myc-” in them. Mycoses are fungal diseases. ascomycetes are a genus of fungi, Mycetophilidae are the fungus – loving gnats. Medical mycologists identify pathogenic fungi by their asexual spores; plant mycologists may deal with fungi that develop perfect states – the fruit that contain sexual spores – and use these for identification. Where perfect states cannot be achieved in cultivation, the structure of the hyphae, the pattern of branching and rejoining of hyphae (anastomosis), or the number of nuclei in cells may give a clue as to identity.
Mycorrhizas
If you read about orchids you can’t escape phrases like “fungal associations”, and “pine needle layer rich in fungal hyphae”. In fact the roots of most vascular plants have evolved in association with soil fungi. The resulting combined structures are called mycorrhizas (“fungus-roots”). There are seven main kinds of mycorrhizas, the four most carefully studied involving crop plants, forest trees, heaths and orchids [1].

Many orchid mycorrhizal fungi belong to the form genera Rhizoctonia, Epulorrhiza, or Ceratorhiza. These same genera may contain species that are orchid pathogens, form associations with other plants, or have no plant associations.
The habitat of the fungus may determine the habitat of the orchid – thus the fungus Rhizoctonia borealis requires acid soils under conifers, so that is where its associated European orchids Spiranthes gracilis and Goodyera repens are found.
Strictly, the term mycorrhiza should apply only to the fungus/root association, but it is loosely association between the fungus and the developing orchid protocorm (the stage between seed and embryo).
Orchid mycorrhizas
Orchids require the relationship with a fungus for their existence. The importance differs among species, the “infection” by the fungus being heaviest in temperate terrestrials, but light in tropical epiphytes. The relationship is essential for the germination of the seed of all orchids in the wild, and remains essential for a few species throughout life.
Seeds, protocorms and fungi

Orchid seeds are tiny and lack the built-in nutrition of bigger seeds; orchids then pass through a nongreen (“achlorophyllous”) developmental stage when they cannot use fats, break down starch, obtain phosphates or photosynthesise, and therefore rely on an external source. This is provided either by man in the form of simple carbon-containing foods in sterile seed germination, or by a fungus which breaks down complex compounds into simpler ones in symbiotic germination. The fungal hyphae penetrate via the base end of the seed. The hyphae enter the cells and coil into structures called pelotons Germination of the seed into a protocorm follows. The cells eventually digest the pelotons, but occasionally the fungi become parasitic and destroy the protocorm.
Roots and fungi
In some species (Gastrodia, Danhatchia and Corybas cryptanthus in New Zealand) chlorophyll never does develop, so the orchids rely for all their lives on associations with fungi. In others, the leaf-size is too small to support the rest of the orchid, and the orchid continues to rely partly on the fungus for its nutrition (Corybas cheesemanii for instance); such plants have been called saprophytic, but that is an incorrect application of the term. Some plants of the European Spiranthes spiralis pass alternate seasons underground, apparently fully nourished by their fungus during that time; some NZ orchids do not appear above ground every year and may do the same.
Most terrestrials seem to thrive better in the wild than in pots (some cannot be cultivated “artificially” at all), probably because they must have access to at least some of their nutrition via their ~ association.
In different terrestrial orchids the fungi penetrate the stems, tubers or root hairs, via epidermal (“skin”) cells after hyphae have spread over the root surface [2]. Pelotons are formed, and eventually digested.
“Symbiosis” suggests mutual benefit and indeed Cymbidium and its fungus each require the vitamin thiamine, made up of thiazole and pyridine; the fungus supplies the thiazole and the orchid supplies the pyridine [3]. Most orchid-associated fungi can, however, live without the orchid, and it seems that whereas the fungus supplies the orchid with a range of nutrients and stimuli, the orchid usually provides little in return.
Many orchids have “host” cells that store fungus, and adjacent digestion cells that break the fungus down by means of substances known as phytoalexins. The partnership between orchid and fungus has been called symbiosis (a ‘Swan situation’ as the politicians say in Wellington these days), or a “delicately balanced mutual antagonism’ [4], or plain parasitism (of the orchid on the fungus, that is.
Fungi that are apparently symbiotic can turn nasty and attack the orchid; furthermore the fungi of epiphytes may invade the orchid’s host tree to the tree’s (and ultimately the orchid’s) detriment.
Specificity
Some studies in the laboratory suggest that specific orchids require specific fungi, but few associations have been studied in the wild. Fungi are difficult to isolate and difficult to grow (especially to the usually readily identifiable perfect state), and even in one orchid species, the fungus required by the protocorm may be different from that required by the adult. Certainly some orchids can establish successful relations with several different fungi.
Perkins has looked at the Australian orchids Pterostylis acuminata and Microtis parviflora in the wild and in the laboratory [19, 20]. Whereas only a few species of fungi were associated in the wild, several more would form associations in the laboratory – thus, ecological specificity” (what happens in the wild) is different from “potential specificity” (what could happen if laboratory experiments were to reflect the wild state) [18].
New Zealand Studies
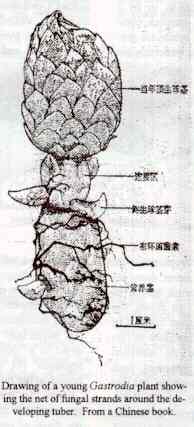
In 1911 Lancaster showed that fungal hyphae do penetrate the root hairs of NZ epiphytes and form pelotons which are digested by the orchid cells [5].
Ella Campbell began a series of papers on the fungal associations of NZ’s nongreen orchids in 1962 [6-10]; she showed: –
- Gastrodia cunninghamii is associated with the fungus Armililaria mellea which is itself a parasite on the roots of forest trees [6].
- G. minor is associated with and derives nutrients from an unidentified fungus which also penetrates the roots of adjacent manuka [7].
- What is probably the bracket fungus Fomes mastopourus inhabits the roots of Acacia melanoxylon and is an endophyte of Gastrodia aff. sesamoides, which digests it [8];
All around the roots of taraire trees grow the hyphae of the puftball fungus Lycoperdon perelatum, and these form a network around and attach to the rhizomes of Danhatchia australis, invade the tips of root hairs, and are digested by the cells of the orchid. The orchid is parasitic on the fungus, which in turn derives nutrients from, and may damage, the roots of the taraire [9].

Corybas cryptanthus has an associated unidentified fungus that invades the roots through root hairs attached at tiny conical projections; the fungus spreads among the beech leaf litter, and is a weak parasite on the Nothofagus[10].
Australian work
J.H. Warcup, M. Clements, K. Dixon and A. Perkins and their co-workers have been the major contributors to the study of Australian orchid/fungus relationships [2, 11-21]. Readers interested in delving deeper are referred to these authors (for instance Warcup [16] gives an excellent general overview of the fungal relationships of South Australian orchids). Here are a few snippets.
- Warcup and Talbot seem to have had a genius for inducing orchid fungi to fruit in culture. They grew fungi from pelotons teased from the cells of Australian native orchids – of 102 isolates from 25 orchid species 66 fungi were induced to fruit. Fungal species of the following genera formed mycorrhizal associations with orchid:: species (of the genera in brackets): Thanatophorous(Acianthus, Thelymitra), Ceratobasidium (Pterostyis, Prasophyllum, Acianthus), Tulasnella (Diuris, Acianthus, Thelymitra, Caladenia, Cymbidium, Dendrobium) and Sebacina (Acianthus, Caladenia, Glossodia, Microtis); the same fungal species often formed mycorrhizal associations with European orchid species. These truly intracellular fungi were often different from those found on the surface of orchid roots [12].
- Fire affected the abundance, behaviour and composition of fungus infecting West Australian orchids; there were six categories of fungus, and each was specific and consistent within species and within most genera; the rare and related Drakaea, Paracaleana and Spiculaea had a unique and culturally distinct fungus noted for its intense violet-pink colour [2].
- Initial contact between fungus and seed is haphazard – there is no evidence that an attractant is used by the orchid seed. Seeds appeared to resist entry by incompatible fungi, while allowing the entry of compatible fungi. There was a strong specificity of fungus for each orchid studied. Pelotons appeared about a week after initial infection in some cells and signified a compatible orchid/fungus match that would lead to germination. The protocorm seemed to have entry, holding and digestion zones for the fungus, though the way the fungus is controlled in these zones is unknown. Failure of germination was caused by fungal hyphae failing to penetrate the seed, or by penetrating all the embryo’s cells resulting in death of the embryo [21].
- In Pterostylis the fungi can be grouped, and where the groups are found is determined by the environment. One fungus, for instance, was found only under Pinus radiata. Geographic distribution (and perhaps some aspects of habitus?) of orchid species may thus be determined by fungal ecology [17].
- Perkins and co-workers found only two fungi associated with Microtis parviflora in the wild, and the same two in protocorms: they concluded that the adult roots associate with a narrow range of fungi in the field (have a narrow ecological specificity) and these assmiations are established in the protocom. On the other hand, many fungi were able to form associations with M. parviflora in the laboratory, indicating a broad potential specificity [19].
- It would seem logical that the germination of orchid seed in the wild should depend on the amount of fungus in the soil, but this may not be so. Perkins and co-workers studied Pterostylis acuminata and its fungal associations: this orchid appears to associate with only one specific fungus, a subspecies of Rhizoctonia solani. Furthermore this orchid reproduces asexually (i.e. essentially by cloning). The orchid and the fungus may therefore be co-distributed, and if an orchid is able to establish at a new site, the resultant increase in the associated fungus may favour further spread of the orchid. There are implications here for the resiting of rare orchids – if there is a single fungus associated with the orchid, a new site will need to be apt for the fungus as well as for the orchid: if the fungus does not survive, neither will the orchid [20].
Orchids that form ecologically specific relationships with single pollinating insects can only survive in the presence of that specific insect. We now see that there are orchids which form ecologically specific relationships with single mycorrhizal fungi: they can only survive in the presence of that specific fungus. How these observations apply to the New Zealand species is open to speculation.
Acknowledgements
I am grateful to Dr A.J. Perkins for supplying a list of Australasian papers.
References
- Peterson RL, Farquhar ML. Mycorrhizas – integrated development between roots and fungi. Mycologia 1994; 86 (3): 311-326.
- Ramsay RR, Dixon KW, Sivasithamparam K. Patterns of infection and endophyte associations with Western Australian orchids. Lindleyana 1986; 1: 203-214.
- Hijner JA, Arditti J. Orchid mycorrhiza; vitamin requirements and production by the symbionts. Amer.J.Bot. 1973; 60: 829-835.
- Arditti J, Fundamentals of orchid biology. Wiley, New York, 1992. p445.
- Lancaster T.L. Preliminary note on the fungi of the New Zealand epiphytic orchids. Trans.N.Z.L 1911; 43: 186-191.
- Campbell E.O. The mycorrhiza of Gastrodia cunninghamii HookF. Trans.Roy.Soc.N.Z 1962; Bot 1: 289.
- Campbell E.O. Gastrodia minor Petrie, an epiparasite of Manuka. Trans.Roy.Soc.N.Z 1963; Bat 2: 73.
- Campbell E.O. The fungal association of a colony of Gastrodia sesamoides R.Br. Trans.Roy.Soc.N.Z. 1964; Bot 2: 237.
- Campbell E.O. The Fungal Association of Yoania australis. Trans.Roy.Soc.N.Z. 1970; Biol.Sci. 12: 5-12.
- Campbell E.O. The Morphology of the Fungal Association of Corybas cryptanthus. J.Roy.Soc.N.Z 1972; 2: 43-47.
- Dixon K. Seeder/clonal concepts in Western Australian orchids. In Population ecology of terrestrial orchids. Eds T.C.E. Wells and J.H. Willems. J.H. SPB Academic Publishing: The Hague, 1991, ppl11-124.
- Warcup J.H. and Talbot P.H.B. Perfect states of Rhizoctonias associated with orchids I-III. New Phytologist 1967; 66: 631-641; 1971; 70: pp35-40; 1980; 86: pp267-272.
- Warcup J. H. Specificity of mycorrhiza association in some Australian orchids. New Phytologist 1971; 70: pp41-46.
- Warcup J. H. Symbiotic germination of some Australian orchids. New Phytologist 1973; 72: pp387-392.
- Warcup J.H. ne mycorrhizal relationship of Australian orchids. New Phytologist 1981; 87: pp371-387.
- Warcup J. H. Mycorrhiza. In Orchids of South Australia. Eds R.J. Bates and J.Z. Weber. Flora and Fauna of South Australia Handbook Committee, Adelaide, 1990. pp21-26.
- Ramsay R.R. Sivasithamparam K. and Dixon K.W. Anastomosis groups among Rhizoctonia-like endophytic fungi in south western Australian Pterostyis species. Lindleana 1987; 2: pp161-167.
- Matsuhara G. and Katsuya K. In situ and in vitro specificity between Rhizoctonia spp. And Spiranthes sinensis. New Phytol. 1994; 127: 711-718.
- Perkins A.J. Masuhara G. McGee P.A. Specificity of the associations between Microtis parviflora and its mycorrhizal fungi Australian J. Bot. 1995; 43: pp85-91.
- Perkins A.J. and McGee P.A Distribution of the orchid mycorhiza1 fungus, Rhizoctonia solani, in relation to its host Pterostyis acuminata, in the field. Australian Journal of Botany 1995; 3(6): pp565-575.
- Clements M. Orchid mycorrhizal associations. Lindleyana 1988; 3: pp73-86.
I notice that all the Reference papers are fairly dated. The most recent three are already 25 years old, these being No 1 and No 18 and No 20 from 1995.
With the explosion of scientific progress in nearly all fields over the last 25 years I would have thought there would be more recent papers referenced.
Hi Martin, You are correct there are new journals that cover some of these aspects. This is old information was about to be lost (wasn’t published in journals) so I was archiving it 🙂 Cheers Tim